This content will be cross-posted to Synthetic Daisies.


This year’s Nobel Prize in Physiology and Medicine went to John O’Keefe, May-Britt Moser, and Edvard I. Moser for their work on the neurophysiology of spatial navigation [1]. The prize was awarded “for their discoveries of cells that constitute a positioning system in the brain”. Some commentators have referred to these discoveries as constituting an “inner GPS system“, although this description is technically and conceptually incorrect (as I will soon highlight). As a PhD student with an interest in spatial cognition, I read (with enthusiasm!) the place cell literature and the first papers on grid cells [2]. So upon hearing they had won, I actually recognized their names and contributions. While recognition of the grid cell discovery might seem to be premature (the original discovery was made in 2005), the creation of iPS cells (the subject of the 2012 award) only dates to 2007.
John O’Keefe is a pioneer in the area of place cells, which provided a sound neurophysiological basis for understanding how spatial cognitive mechanisms are tied to their environmental context. The Mosers [3] went a step further with this framework, discovering a type of cell that provides the basis for a metric space (or perhaps more accurately, a tiling) to which place cell and other location-specific information are tied. The intersection points on this grid are represented by the aptly-named grid cells. Together, these types of cells provide a mental model of the external world in the medial temporal lobe of mammals.
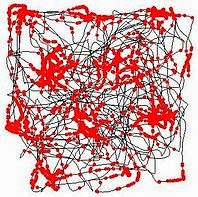
Locations to which grid cells respond most strongly.
Place cells (of which there are several different types) are small cell populations in the CA1 and CA3 fields of the Hippocampus that encode a memory for the location of objects [4]. Place cells have receptive fields which represent specific locations in space. In this case, a cell’s receptive field corresponds to locations and orientations to which the cell responds most strongly. When the organism is located in (or approaches) one of these receptive fields, the local field potential of the cell population is activated at a maximum of 20Hz. As place cells are in the memory encoding center of the brain, place cells respond vigorously when an animal passes or gets near a recognized location. Grid cells, located in the entorhinal cortex, serve a distinct but related role to that of place cells. While spatial cognition involves many different types of input (from multisensory to attentional), place cells and grid cells are specialized as a mechanism for location-specific memory.

Variations on a grid in urban layouts. COURTESY: Athenee: the rise and fall of automobile culture.
How do we know this part of the brain is responsible for such representations. Both place and grid cells have been confirmed through electrophysiological recordings. In the case of place cells, lesions studies [5] have been conducted to demonstrate behavioral deficits during naturalistic behavior. In [5], lesions (made via lesion studies) of hippocampal tissue results in deficits in spatial memory and exploratory behavior. In humans, the virtual Morris Water Maze [6] can be used to assess performance with regard to finding a specific landmark (in this case, a partially-submerged platform) embedded in a virtual scene. The recall of a particular location is contingent on people’s ability to a) find a location relative to other landmarks, and b) people’s ability to successfully rotate their mental model of a particular space.

An example of learning in rats during the Morris Water Maze task. COURTESY: Nutrition, Neurogenesis and Mental Health Laboratory, King’s College London.
As a relatively recent discovery, grid cells provide a framework for a geometric (e.g. Euclidean) representation of space. Like place cells, the activity of grid cells are dependent upon the behavior of animals in a spatial context. Yet grid cells help to provide a larger context for spatial behavior, namely the interstitial space between landmarks. This allows for both the creation and recognition of patterns at the landscape spatial scale. Street patterns in urban settlements that form grids and wheel-and-spoke patterns are no accident — it is the default way in which humans organize space.
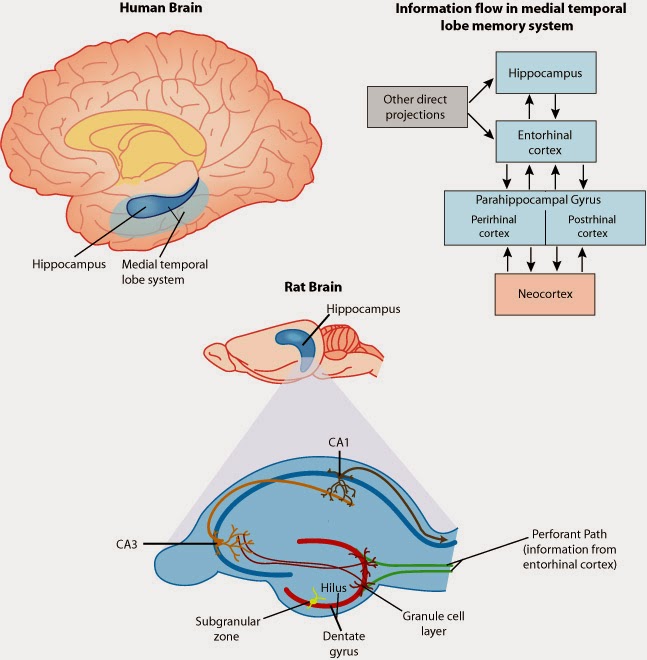
An anatomical and functional view of the medial temporal lobe. COURTESY: Figure 1 in [7].
There are some interesting but unexplored relationships between physical movement and spatial navigation which both involve a coordinate system for the world that surrounds a given organism. For example, goal-directed arm movements occur within a multimodal spatial reference frame that involves the coordination of visual and touch information [8]. While limb movement and walking involve timing mechanisms associated with the motor cortex and cerebellum, there are implicit aspects of spatial memory in movement, particularly over long distances and periods of time. There is an emerging field called movement ecology [9] which deals with these complex interconnections.
Another topic that falls into this intersection is path integration [10]. Like the functions that involve place and grid cells, path integration also involves the medial temporal lobe. Path integration is the homing ability of animals that results from an odometer function — the brain keeps track of footsteps and angular turns in order to generate an abstract map of the environment. This information is then used to return to a nest or home territory. Path integration has been the basis for digital evolution studies on the evolutionary origins of spatial cognition [11], and might be more generally useful in understanding the relationships between the evolutionary conservation of spatial memory and its deployment in virtual environments and city streets. While this is closer to the definition of an “inner GPS system”, there is so much more to this fascinating neurophysiological system.
NOTES:
[1] Nobel Prize Committee: The Nobel Prize in Physiology or Medicine 2014. Nobelprize.org, Nobel Media AB. October 6 (2014).
[2] Hafting, T., Fyhn, M., Molden, S., Moser, M-B., and Moser, E.I. Microstructure of a spatial map in the entorhinal cortex. Nature, 436(7052), 801–806 (2005).
[3] Moser, E.I., Kropff, E., and Moser, M-B. Place Cells, Grid Cells, and the Brain’s Spatial Representation System. Annual Review of Neuroscience, 31, 69-89 (2008).
[4] O’Keefe, J. and Nadel, L. The Hippocampus as a Cognitive Map. Oxford University Press (1978).
[5] For the original Morris Water Maze paper: O’Keefe, R.G., Garrud, P., Rawlins, J.N., and O’Keefe, J. Place navigation impaired in rats with hippocampal lesions. Nature, 297(5868), 681–683 (1982).
[6] For the virtual adaptation of the water maze for humans, please see: Astur, R.S., Taylor, L.B., Mamelak, A.N,, Philpott, L., and Sutherland, R.J. Humans with hippocampus damage display severe spatial memory impairments in a virtual Morris water task. Behavioral Brain Research, 132, 77–84 (2002).
[7] Bizon, J.L. and Gallagher, M. More is less: neurogenesis and age-related cognitive decline in Long-Evans rats. Science of Aging, Knowledge, and Environment, (7), re2 (2005).
[8] Shadmehr, R. and Wise, S.P. The Computational Neurobiology of Reaching and Pointing. MIT Press, Cambridge, MA (2005).
[9] Nathan, R. An emerging movement ecology paradigm. PNAS, 105(49), 19050–19051 (2008).
[10] McNaughton, B.L., Battaglia, F.P., Jensen, O., Moser, E.I., and Moser, M-B. Path integration and the neural basis of the ‘cognitive map’. Nature Reviews Neuroscience, 7, 663-678 (2006).